|
In 1925 Dutch Physiologists Gorter and Grendel pproached the discovery of our present model of the plasma membrane structure as a lipid bi-layer with the polar hydrophilic heads facing outwards towards the aqueous environment and the hydrophobic tails facing inwards away from the aqueous surroundings on both sides of the membrane. In 1935 physiologists Davson and biologist Danielli refined the Gorter-Grendel model adding to the phospholipds bi-layer a proteins mono-layer. 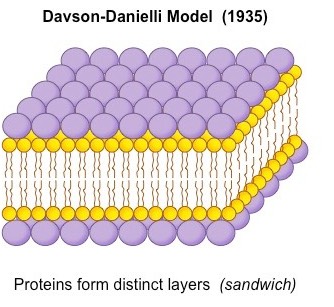 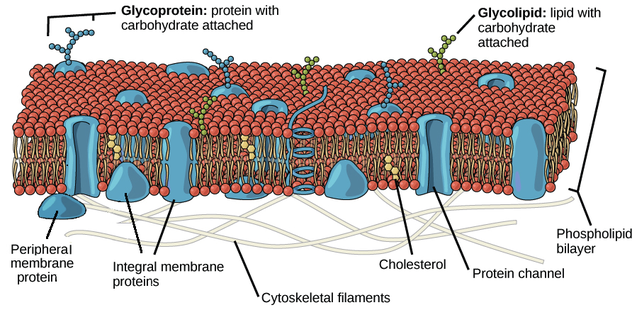 Reasons for the model:
1 | Chemical analysis of membranes showed that they were composed of phospholipid and protein 2 | Evidence suggested that the plasma membrane of red blood cells has enough phospholipids in it to form an area twice as large as the area of the plasma membrane, suggesting a phospholipid bi-layer. 3 | Experiments showed that membranes form a barrier to the passage of some substances, despite being very thin, and layers of protein could act as the barrier. Testing the model In the 1950s, thanks to high magnification micrographs, Davson and Danielli could have proof of their theory: in these in fact, membranes appeared as two dark lines that seemed proteins separated by a lighter band of possibly phospholipids. In 1960s evidence accumulated that did not fit the Davson-Danielli model: 1 | Freeze-fracture electron micrographs showed that globular proteins were present in the centre of the phospholipid bilayer 2 | Analysis of membrane proteins showed that parts of their surfaces were hydrophobic, so they would be positioned in the bilayer and in some cases would extend from one side to the other 3 | Fusion of cells with membrane proteins tagged with different coloured fluorescent markers showed that these proteins can move within the membrane as the colours became mixed within a few minutes of cell fusion After the Davson-Danielli model was falsified, in 1966, a new membrane model was proposed by American biologists Singer and Nicolson They firstly described the cell membrane as selectively permeable or only allowing certain substances to pass through it. Its composition was therefore best described by the fluid mosaic model, where different parts of the membrane can float in a fluid-like space. Like it sounds, this model suggests that the cell membrane is dynamic or moving and asymmetrically scattered with different pieces, such as phospholipids, cholesterol, proteins, and carbohydrates, and that these different parts of the cell membrane float in this fluid-like space, moving laterally throughout the membrane. PHOSPHOLIPIDS Phospholipids are the basic component of all biological membranes. Phospholipid molecules are amphipathic. This means part of the molecule, the phosphate head is attracted to water ( hydrophilic ) and part, made of the two fatty acid tails composed by hydrocarbon chains, is not attracted to water. The fatty acid hydrophobic tails of the phospholipids, face each other, creating this hydrophobic region inside the bilayer. A function of this bilayer is to create a distinct hydrophobic region which is essential in both keeping fluidity and moderating who can get by the membrane. CHOLESTEROL Cholesterol is a component of all animal cell membranes. Most of its molecule is hydrophobic but, like phospholipids, there is one hydrophilic end; so cholesterol fits between phospholipids in the membrane. Cholesterol restricts the movement of phospholipid molecules. It therefore reduces the fluidity of the membrane, as well as its permeability to hydrophilic particles such as sodium ions and hydrogen ions. This is important, as animal cells need to maintain concentration differences of these ions across their membranes, so diffusion through the membrane must be restricted. MEMBRANE PROTEINS Classifications of Membrane Proteins Proteins are generally broken down into the smaller classifications of integral proteins, peripheral proteins, and lipid-bound proteins. Integral Proteins Integral proteins are embedded within the lipid bilayer. They cannot easily be removed from the cell membrane without the use of harsh detergents that destroy the lipid bilayer. Integral proteins float rather freely within the bilayer, much like oceans in the sea. In addition, integral proteins are usually transmembrane proteins, extending through the lipid bilayer so that one end contacts the interior of the cell and the other touches the exterior. The stretch of the integral protein within the hydrophobic interior of the bilayer is also hydrophobic, made up of non-polar amino acids. Like the lipid bilayer, the exposed ends of the integral protein are hydrophilic. When a protein crosses the lipid bilayer it adopts an alpha-helical configuration. Transmembrane proteins can either cross the lipid bilayer one or multiple times. The former are referred to as single-pass proteins and the later as multi-pass proteins. As a result of their structure, transmembrane proteins are the only class of proteins that can perform functions both inside and outside of the cell. Peripheral Proteins Peripheral proteins are attached to the exterior of the lipid bilayer. They are easily separable from the lipid bilayer, able to be removed without harming the bilayer in any way. Peripheral proteins are less mobile within the lipid bilayer. Lipid-Bound Proteins Lipid-bound proteins are located entirely within the boundaries of the lipid bilayer.
2 Comments
PRINCE K
5/17/2018 04:54:30 am
Atleast i have got something compared to what i got in class
Oscar
10/29/2018 05:25:55 pm
who wrote this and when was it published? needed for apa citation Leave a Reply. |
ABOUT
IB Biology student at Ardingly College |
 RSS Feed
RSS Feed
